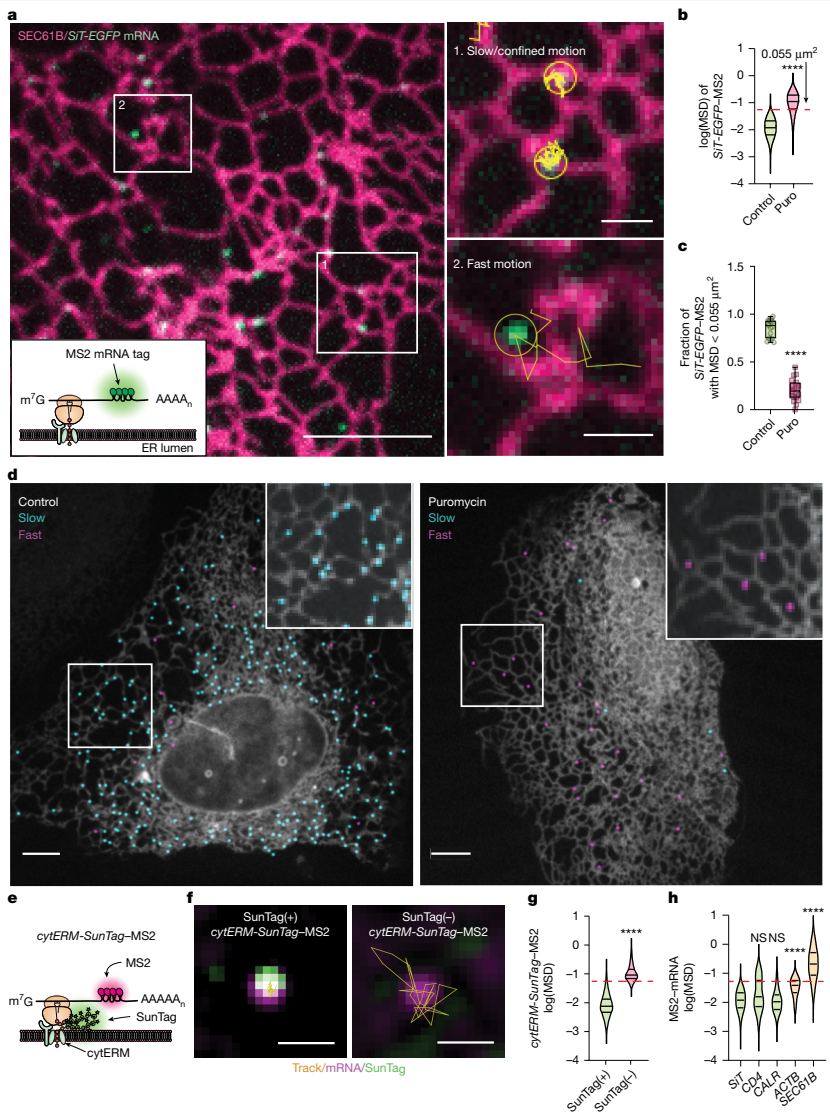
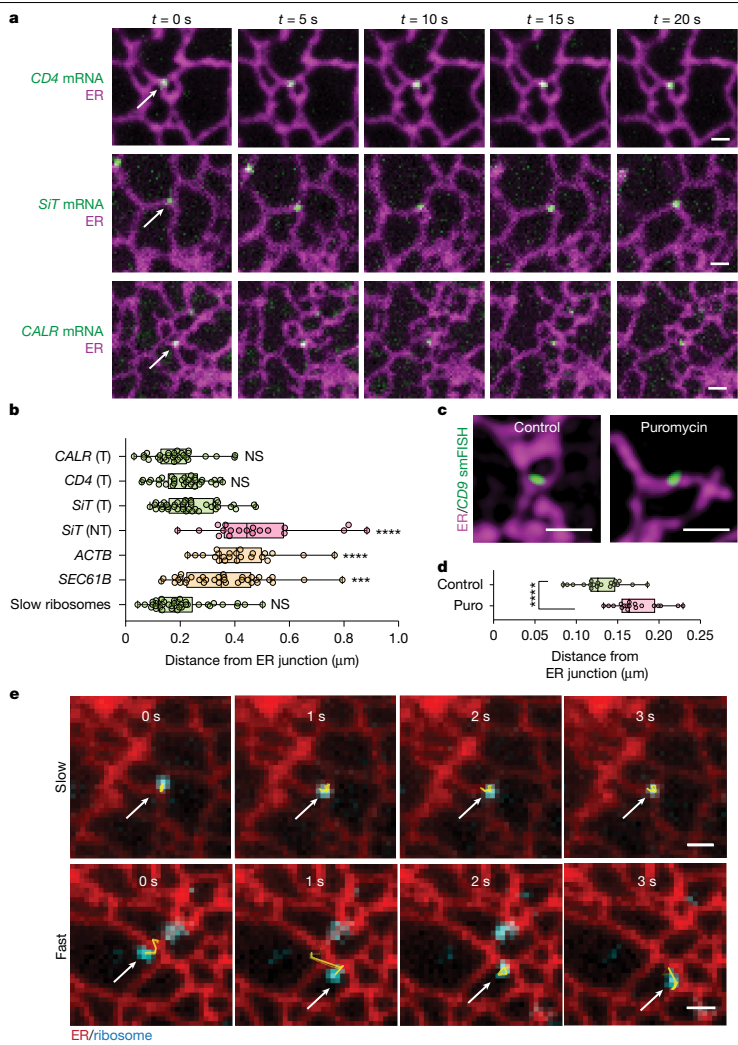
在细胞生物学领域,内质网作为蛋白质合成的重要场所,其复杂的三维网络结构一直备受关注。传统观点认为,分泌组mRNA(编码分泌蛋白、腔内蛋白和整合膜蛋白的mRNA)的翻译主要发生在ER片层结构上。然而,近年来的研究发现,核糖体实际上分布在整个ER网络的各个形态区域,包括小管和管状连接点。更令人困惑的是,有相当一部分与ER结合的核糖体处于非翻译状态,这提示我们,核糖体的结合并不等同于活跃的延伸过程,可能反映了翻译的调控或预起始状态。一个关键的科学问题由此产生:分泌组mRNA的翻译是随机分布在连续的ER网络上,还是存在特定的空间组织?如果存在空间特异性,又是哪些细胞器结构和分子机制在调控这一过程?解答这些问题对于理解细胞如何精确协调蛋白质合成与后续的加工、修饰及转运过程至关重要。为了深入探究这一科学问题,Heejun Choi等研究人员在《Nature》杂志上发表了题为"Secretome translation shaped by lysosomes and lunapark-marked ER junctions"的重要研究。该研究通过创新的活细胞单分子成像技术,首次揭示了分泌组mRNA翻译的空间组织规律,并阐明了LNPK标记的ER连接点与溶酶体在这一过程中的协同调控作用。研究人员主要运用了以下几种关键技术方法:首先,他们开发了多种MS2和SunTag标记的分泌组mRNA报告系统,通过单粒子追踪技术实时监测mRNA的动力学行为;其次,利用杂交链反应单分子荧光原位杂交(HCR-smFISH)技术对内源性CD9 mRNA进行定位分析;第三,通过光遗传学招募实验研究ER-溶酶体接触的形成动力学;此外,还结合了免疫荧光、邻近连接 assay(PLA)、荧光恢复 after photobleaching(FRAP)以及RNA测序和质谱分析等多种技术手段,从多个维度验证了研究结论。ER连接点作为翻译热点通过单分子追踪技术,研究人员发现分泌组mRNA在ER上表现出两种截然不同的运动模式:一部分mRNA保持相对静止或局限在特定区域,而另一部分则快速移动。进一步分析表明,这种运动差异与mRNA的翻译状态直接相关——处于活跃翻译状态的mRNA表现出高度局限的运动特性(MSD<0.055μm2),而非翻译状态的mRNA则运动迅速。
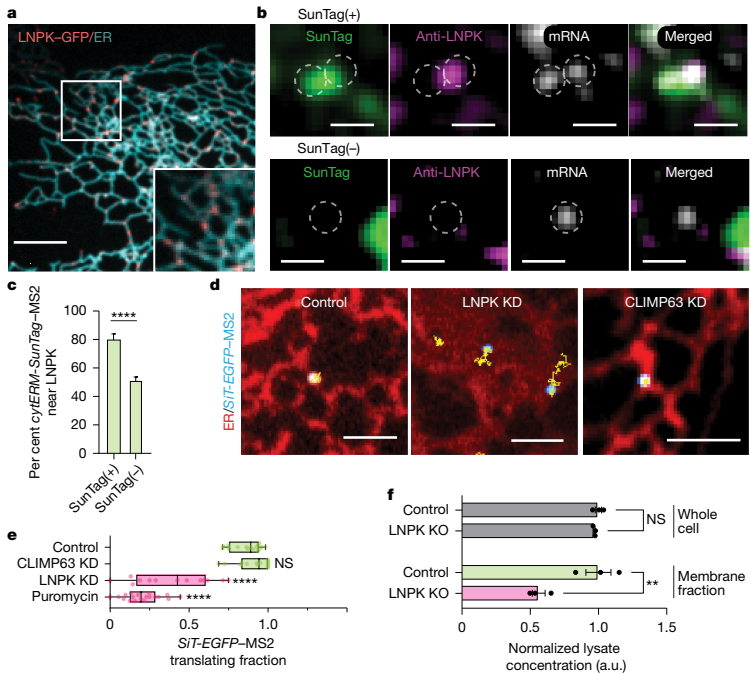
更重要的是,研究人员发现活跃翻译的分泌组mRNA显著富集在ER连接点处。这种连接点偏好性在多种细胞系(U-2 OS、HeLa、COS-7和HT1080)中均得到验证,且通过内源性CD9 mRNA的定位研究进一步证实了这一现象。核糖体追踪实验也显示,只有慢速运动的核糖体群体定位于ER连接点,其运动特性与翻译中的分泌组mRNA高度一致。LNPK阳性连接点的翻译功能ER连接点的形成和稳定依赖于ER驻留跨膜蛋白LNPK。研究发现,LNPK-GFP特异性地定位于ER连接点,而活跃翻译的分泌组mRNA与LNPK信号表现出强烈的共定位。定量分析显示,80%的翻译中mRNA位于LNPK信号300纳米范围内,而非翻译mRNA仅有50%处于此区域。当研究人员敲低LNPK表达后,分泌组mRNA的翻译分数从86%显著下降至55%,而ER形态蛋白CLIMP63的敲低则无此效应。LNPK敲除细胞中,膜蛋白水平显著降低,但总蛋白合成未受影响,质谱分析进一步证实许多膜相关基因的蛋白质丰度 per mRNA 明显下降。
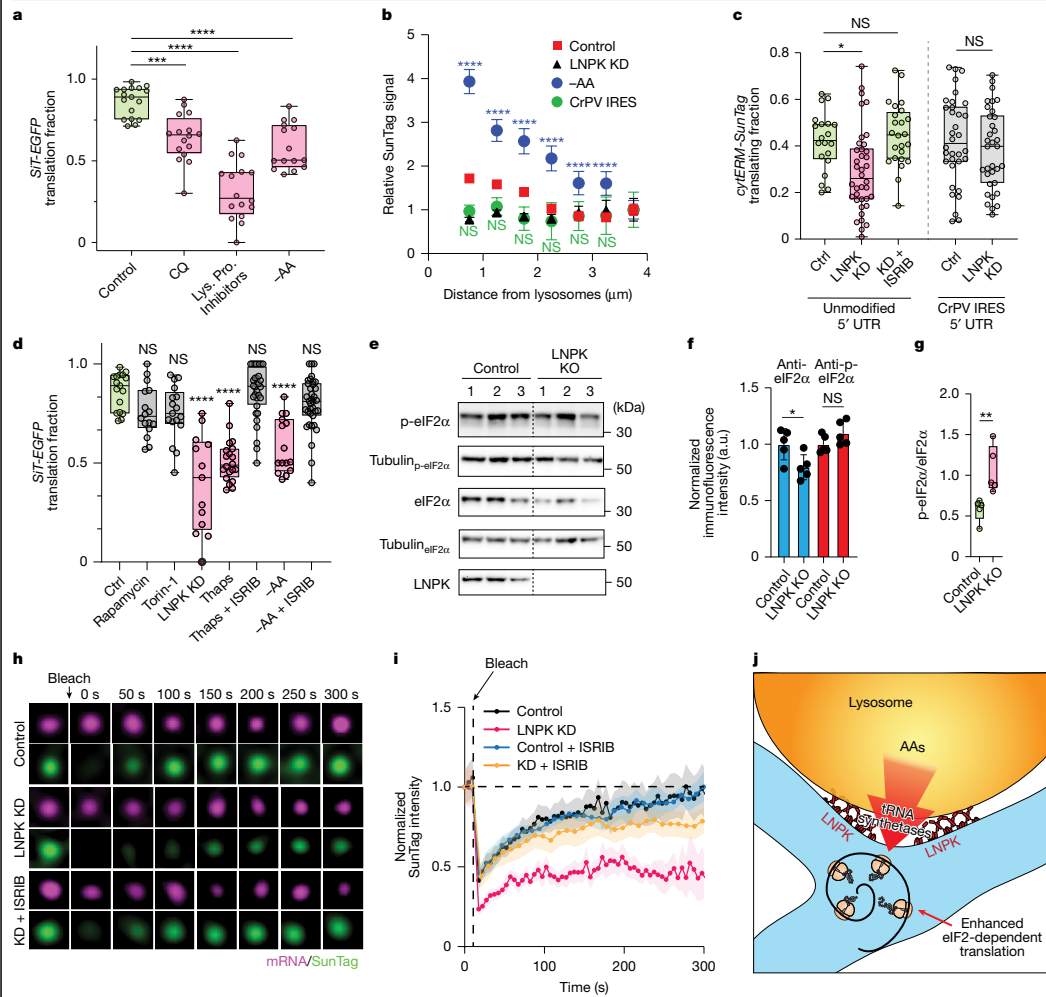
ISRIB恢复LNPK敲低后的翻译通过SunTag斑点的荧光恢复 after photobleaching(FRAP)实验,研究人员直接监测了分泌组mRNA的新生肽链合成。在对照细胞中,光漂白后300秒内荧光可完全恢复至前水平,而LNPK敲低细胞的恢复动力学显著受损。值得注意的是,ISRIB处理可恢复LNPK敲低细胞的SunTag恢复动力学至对照水平,进一步证实了eIF2依赖性机制在LNPK调控分泌组翻译中的核心作用。