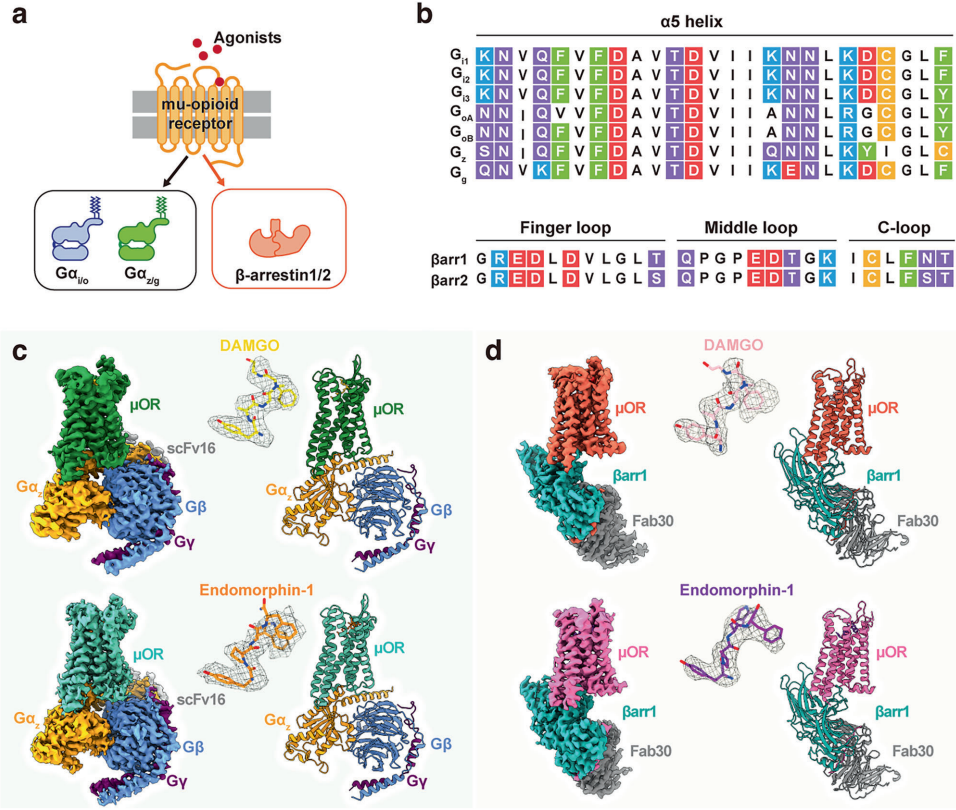
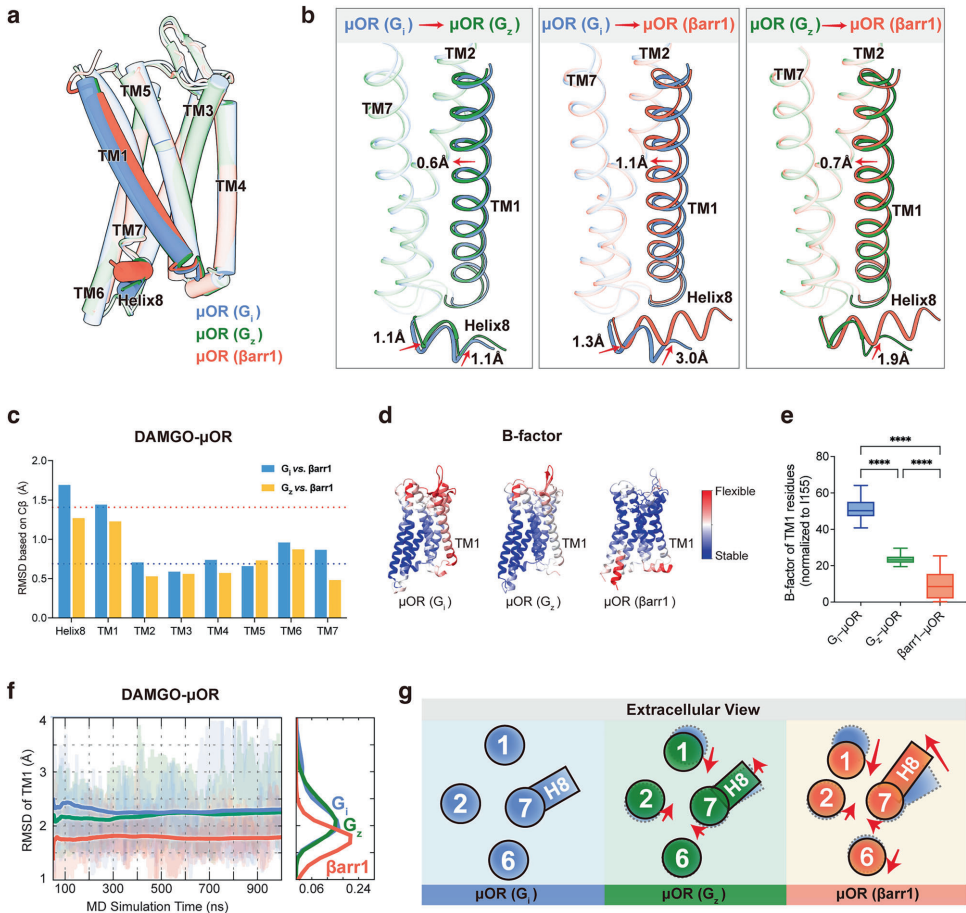
z and μOR-Barr1 complexes. a Schematic diagram of the downstream signal transduction of μOR. b Sequence alignment of the Gαi/o family α5 helices and the βarr1/2 finger loop, middle loop, and C-loop. c Cryo-EM density maps and cartoon representations of the uOR-G2-scFv16 complexes colored by subunit. d Cryo-EM density maps and cartoon representations of the uOR-βarr1-Fab30 complexes colored by subunit.'>Structures of the μOR-Gz and μOR-βarr1 complexes研究团队成功解析了DAMGO和endomorphin-1两种激动剂激活的μOR-Gz和μOR-βarr1复合物结构,分辨率均达2.8 Å。这些结构代表了目前GPCR-β-抑制蛋白复合物的最高分辨率结构,为理解μOR信号转导提供了精确的结构框架。结构比较显示,与Gi结合的μOR相比,Gz和βarr1结合的受体表现出独特的构象特征,特别是在TM1和helix8区域。TM1 and helix8 dynamics in various adaptor couplings of μOR通过比较Gi、Gz和βarr1结合的μOR结构,研究发现TM1和helix8的构象变化与下游信号蛋白的选择性密切相关。在βarr1结合的μOR中,TM1向TM2/TM7方向内移1.1 Å,helix8上移3.0 Å,这种构象变化稳定了受体胞内结合口袋,有利于β-抑制蛋白的招募。B因子分析和分子动力学模拟进一步证实,TM1在βarr1复合物中具有更高的刚性,而在G蛋白复合物中则更灵活。
z-coupled μOR, and Gi-coupled μOR activated by DAMGO. c RMSD comparison of TMs 1-7 and helix8 of μOR. d Gi-, Gz-, and Barr1-coupled μOR activated by DAMGO, colored by B-factors.'>TM1 regulates μOR's downstream signaling selectivity研究人员通过系统突变TM1上与TM2-TM7-helix8界面相互作用的关键残基,发现这些突变对不同信号通路产生差异化影响。例如,Y751.39F突变几乎完全消除βarr1信号而保留G蛋白信号;N861.50A突变显著抑制βarr1招募,但增强Gz信号传导。这些结果表明TM1通过其与受体其他区域的相互作用网络,精细调控下游信号蛋白的选择性招募。
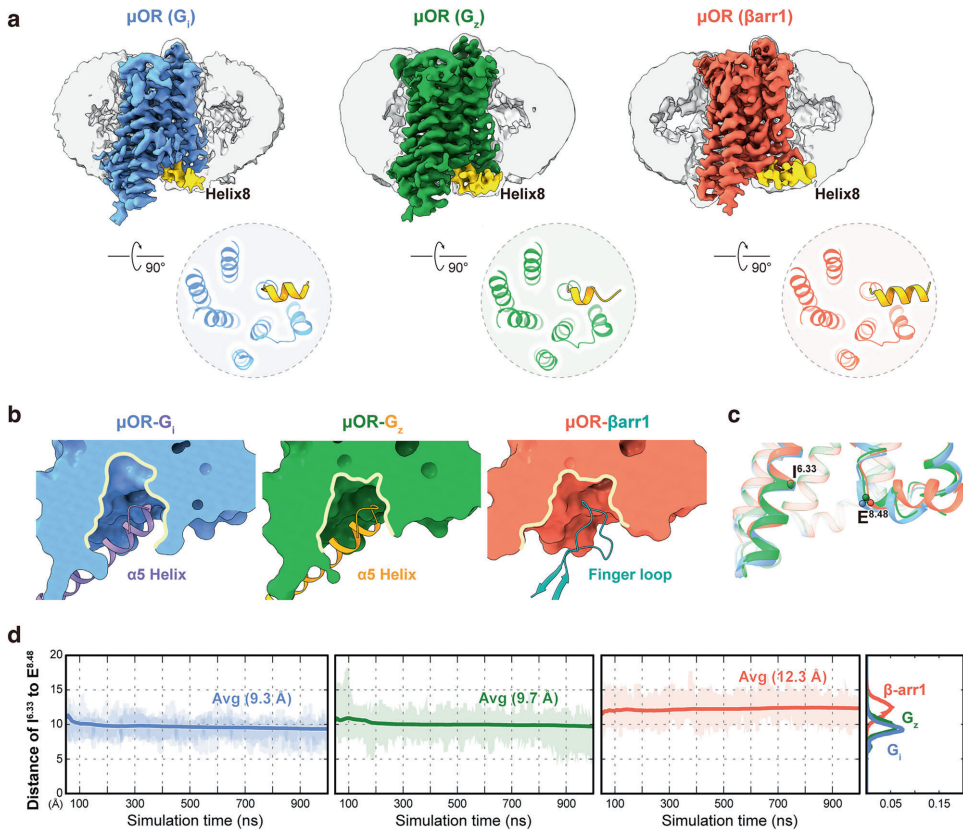
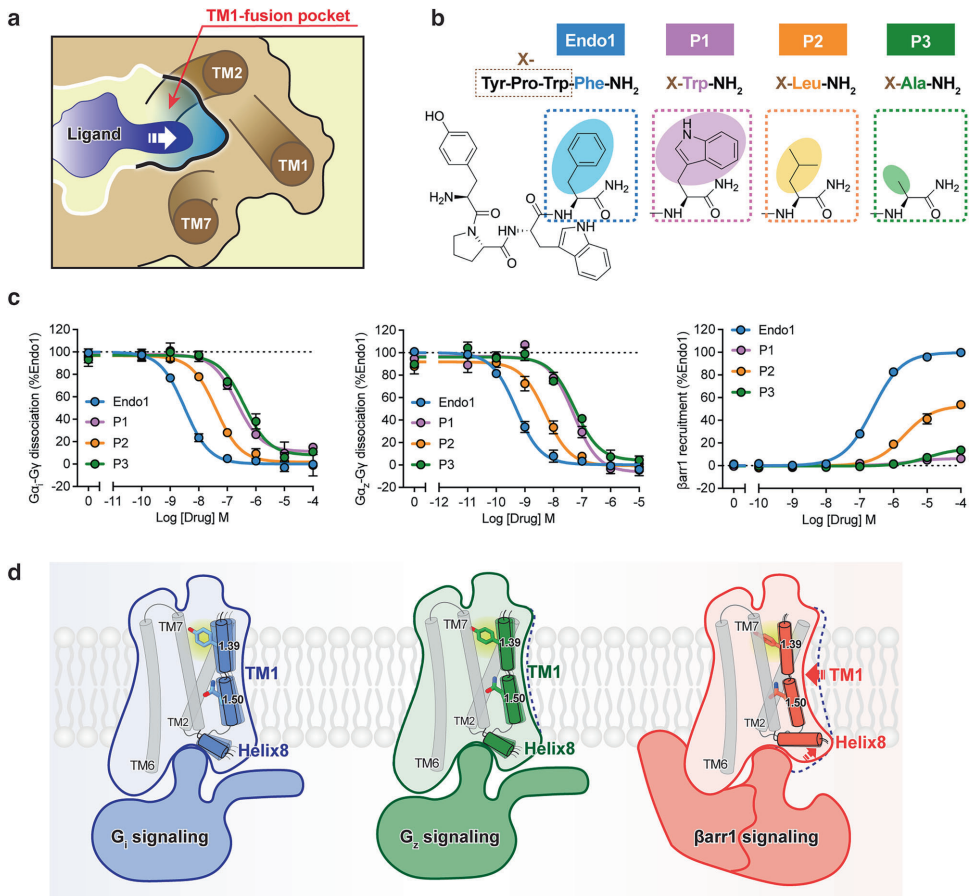
i, Gz, and Barr1. b Comparison of the intracellular halves of the μOR-Gi, μOR-Gz and μOR-βarr1 complexes. c Superposed intracellular structures of μOR coupled with Gi, Gz, and Barr1.'>Propagation of μOR conformational changes研究揭示了μOR激活过程中的构象传播机制:激动剂结合导致胞外区域构象变化,通过保守极性网络(Q2.60D3.32Y7.43)和分子开关W2936.48将信号传递至胞内区域,诱导TM6外移和胞内口袋开放。不同信号蛋白的结合进一步调节TM1和helix8的构象,这些变化反向传播至胞外区域,形成完整的变构调控回路。Activating μOR with DAMGO and endomorphin-1比较两种激动剂激活的μOR复合物发现,endomorphin-1的Phe4侧链插入由TMs 1/2/7形成的TM1融合口袋,而DAMGO则通过其肽链中间区域与TM2-TM7界面相互作用。这种结合模式的差异导致两种激动剂在调控TM1构象和下游信号偏向性方面表现出不同特性。The mechanism of biased signal transduction in μOR基于结构洞察,研究人员设计了一系列endomorphin-1变体肽。实验表明,将Phe4突变为色氨酸(P1)或丙氨酸(P3)可消除βarr1信号而保留部分G蛋白信号,证实通过调控TM1融合口袋可实现对信号通路选择性的精确调控。
i-, Gz-, and Barr1-mediated μOR signaling.'>本研究通过高分辨率结构生物学方法结合功能实验,系统阐明了μOR信号转导的可塑性机制。研究发现TM1作为关键的构象调节器,通过其动态位移变构调控受体对G蛋白和β-抑制蛋白的选择性:TM1向外位移增强G蛋白招募,而向内收缩则促进β-抑制蛋白结合。这一机制的核心在于TM1构象变化调节TM2-TM7-helix8界面的稳定性,进而影响胞内结合口袋的形状和大小。该研究的意义在于首次揭示了TM1在GPCR信号偏向性中的核心作用,挑战了传统上重点关注TMs 5-7的固有认知。鉴定出的TM1融合口袋为开发偏向性μOR激动剂提供了新靶点,有望推动设计具有更低副作用的镇痛药物。此外,TM1的调控机制可能普遍适用于其他GPCRs,为理解这一重要受体家族的信号转导多样性提供了新范式。