编辑推荐:
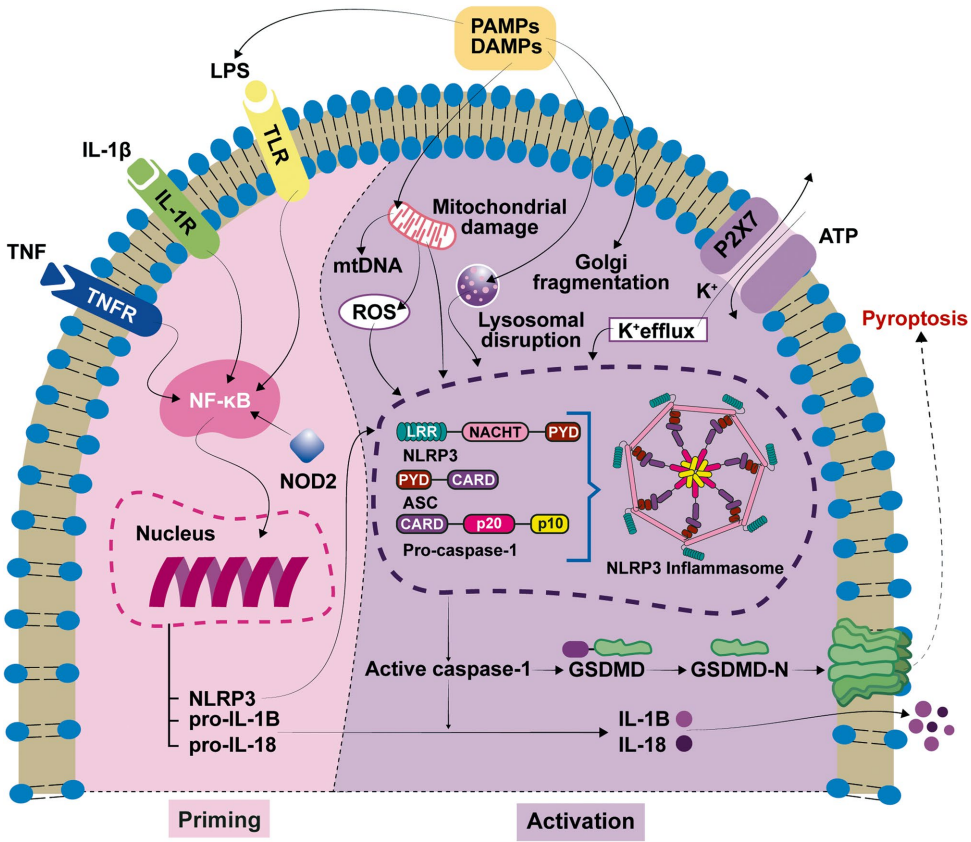
本综述系统总结了NLRP3炎症小体作为先天免疫系统的关键组分,在多种感觉神经性听力损失(SNHL)中的核心作用。文章详细阐述了其在遗传综合征(如CAPS和DFNA34)及噪声性、年龄相关性、药物性和病毒性听力损失中的激活机制,指出异常活化会导致促炎细胞因子(IL-1β、IL-18)释放和细胞焦亡,造成不可逆的耳蜗损伤。文章重点讨论了靶向NLRP3(如MCC950、oridonin)及IL-1β信号通路(如anakinra、canakinumab)的治疗策略在临床前及临床研究中的巨大潜力,为其作为治疗SNHL的新靶点提供了有力证据。
生物通 版权所有





